|
http://www.plosone.org/article/info:doi/10.1371/journal.pone.0006564
William T. Silkworth1, Isaac K. Nardi1, Lindsey M. Scholl2, Daniela Cimini1*
1 Virginia Tech, Department of Biological Sciences, Blacksburg, Virginia, United States of America, 2 Department of Biology, Oberlin College, Oberlin, Ohio, United States of America
Many cancer cells display a CIN (Chromosome Instability) phenotype, by which they exhibit high rates of chromosome loss or gain at each cell cycle. Over the years, a number of different mechanisms, including mitotic spindle multipolarity, cytokinesis failure, and merotelic kinetochore orientation, have been proposed as causes of CIN. However, a comprehensive theory of how CIN is perpetuated is still lacking. We used CIN colorectal cancer cells as a model system to investigate the possible cellular mechanism(s) underlying CIN. We found that CIN cells frequently assembled multipolar spindles in early mitosis. However, multipolar anaphase cells were very rare, and live-cell experiments showed that almost all CIN cells divided in a bipolar fashion. Moreover, fixed-cell analysis showed high frequencies of merotelically attached lagging chromosomes in bipolar anaphase CIN cells, and higher frequencies of merotelic attachments in multipolar vs. bipolar prometaphases. Finally, we found that multipolar CIN prometaphases typically possessed γ-tubulin at all spindle poles, and that a significant fraction of bipolar metaphase/early anaphase CIN cells possessed more than one centrosome at a single spindle pole. Taken together, our data suggest a model by which merotelic kinetochore attachments can easily be established in multipolar prometaphases. Most of these multipolar prometaphase cells would then bi-polarize before anaphase onset, and the residual merotelic attachments would produce chromosome mis-segregation due to anaphase lagging chromosomes. We propose this spindle pole coalescence mechanism as a major contributor to chromosome instability in cancer cells.
Citation: Silkworth WT, Nardi IK, Scholl LM, Cimini D (2009) Multipolar Spindle Pole Coalescence Is a Major Source of Kinetochore Mis-Attachment and Chromosome Mis-Segregation in Cancer Cells. PLoS ONE 4(8): e6564. doi:10.1371/journal.pone.0006564
Editor: Kevin G. Hardwick, University of Edinburgh, United Kingdom
Received: May 10, 2009; Accepted: July 3, 2009; Published: August 10, 2009
Copyright: © 2009 Silkworth et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: IKN was a recipient of a Fralin Summer Undergraduate Research Fellowship in summer 2008 and summer 2009. This work was partially supported by NSF grant MCB-0842551 and the Thomas F. and Kate Miller Jeffress Memorial Trust grant J-828 to DC. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: The authors have declared that no competing interests exist.
* E-mail: cimini@vt.edu
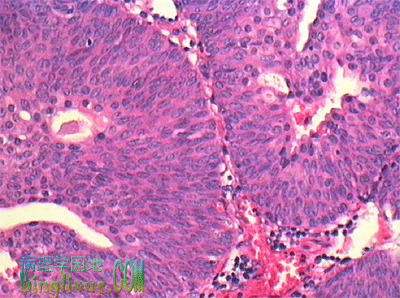
移行细胞癌肾盂、输尿管、膀胱粘膜上皮及尿道的部分上皮均为移行上皮,最常发生的肿瘤为移行细胞癌。鳞癌和腺癌也偶有发生。移行细胞癌多发生在膀胱,有多中心性发生倾向,提示致癌因子可能在尿中。发病相关因素包括吸烟和接触化学性致癌物(纺织、印刷所用的染料,橡胶、电缆、塑料工业所用的试剂等)。 男性多于女性(男性是女性的3倍),高龄人群(60-70岁)多发。始发症状,血尿占70%~90%,其中60%~70%为无症状性血尿。部分病例是在门诊检查时无意中发现的。未侵及粘膜下的浅表性膀胱癌预后较好,五年生存率60%~100%。但浸润性膀胱癌进展迅速,五年生存率仅25~60%。血行转移多及肺、肝和骨,淋巴道转移多见于腹膜后和主动脉周围的淋巴结。病理变化移行细胞癌的好发部位为膀胱侧壁和三角区近输尿管开口处,单发或多发,大小不等(数mm~数cm)。呈乳头状、息肉状或扁平的突起状,并向深层浸润。切面灰白,可有坏死等改变。根据细胞分化程度可将移行细胞癌分为三级。Ⅰ级:呈乳头状生长。癌细胞具有明显的极性,多层(可超过7层),似移行上皮,细胞仅见轻微异型性,核异型和核分裂像罕见;Ⅱ级,肿瘤细胞增生达7层以上,仍保留乳头状增生的形态,但细胞的极性部分紊乱,从基底部向表层分化倾向不明了,可见大且深染的异型核,茎部和粘膜下浸润;Ⅲ级,癌细胞呈巢状分布,乳头状增生倾向不明显,癌细胞异型性明显,核大,大小不一,深染,核分裂像多,癌巢中央细胞坏死明显,肿瘤边缘常侵及粘膜下层和肌层。预后受组织学分级,有无血管、淋巴管浸润,膀胱壁内的浸润深度,有无转移等因素影响。http://www.binglixue.com/lilun/ren41.htm
 |
肿瘤的异型性( atypia)由于分化的程度不同,肿瘤的细胞形态和组织结构与相应的正常组织相比,有不同程度的差异,病理学将这种差异成为异型性。
有两个方面:细胞异型性和结构异型性。 (一)肿瘤的细胞异型性(cellular atypia)有以下表现:(1)肿瘤细胞通常比相应正常细胞大;(2)肿瘤细胞的大小和形态很不一致(多形性),可以出现瘤巨细胞,即体积很大的肿瘤细胞。但是,有些分化甚差的肿瘤,其瘤细胞很原始,体积不大,大小和形态也可以比较一致;(3)肿瘤细胞核的体积增大。胞核与细胞浆的比例(核浆比)增高。例如,上皮细胞的核浆比正常时多为1:4~1:6,恶性肿瘤细胞则可为1:1;(4)核的大小、形状和染色差别较大(核的多形性)。可出现巨核、双核、多核或奇异形的核。核内DNA常增多,核深染(hyperchromasia),染色质呈粗颗粒状,分布不均匀,常堆积在核膜下;(5)核仁明显,体积大,数目也可增多;(6)核分裂像常增多,出现病理性核分裂像,如不对称核分裂、多极性核分裂等。
良性肿瘤细胞一般异型性较小,恶性肿瘤细胞则常具有高度异型性。上述瘤细胞的形态,特别是胞核的多形性和病理性核分裂,常为恶性肿瘤的重要特征,在区别良恶性肿瘤上有重要意义。异型性越大,表示肿瘤组织和细胞与相应正常组织的差异越大。异型性是肿瘤组织和细胞出现成熟障碍和分化障碍的表现。一般来说,异型性越大,成熟程度和分化程度就越低。有些恶性肿瘤细胞分化很差,异型性显著,称为间变性肿瘤(anaplastic neoplasm)。间变性的肿瘤细胞常具有多形性(pleomorphism),即肿瘤细胞的大小和形状变异很大。间变性肿瘤多为高度恶性的肿瘤。
(二)肿瘤组织在空间排列方式上与相应正常组织的差异称为肿瘤的结构异型性(architectural atypia)。如食道鳞状细胞癌中,鳞状上皮排列的极向显著紊乱;胃腺癌中腺上皮失去极向,形成的腺体很不规则;子宫内膜腺癌中,腺体之间正常的内膜间质消失等等。良性肿瘤的细胞异型性一般较小,但可有不同程度的结构异型性。恶性肿瘤的细胞异型性和结构异型性都比较明显。
http://baike.baidu.com/view/1039007.htm
肿瘤细胞染色体不稳定的机制Nature 460 (7252)
发表日期:2009年7月9日
很多肿瘤细胞的一个显著特征是染色体不稳定(CIN),即全部染色体数量的增加或减少速度加快。CIN的机制一直是一个有很多争论的话题。人们长期所持的一个观点是,额外的中心体(它们与CIN有关联性)通过诱导混乱的多极纺锤体组装和非对称的细胞分裂来促使染色体错误分离。但是现在,长期活细胞成像显示,CIN癌细胞很少经历多极分裂,而且甚至当它们经历这种分裂的时候,所产生的也主要是无法生存的后代。相反,额外中心体癌细胞仅仅是经过一个过渡性的多极纺锤体阶段,在这个阶段,“异常动粒微管附着体”会积累。很多附着错误即使在额外中心体聚集形成正确双极纺锤体之后仍存在,从而增加染色体错误分离的可能性。本期封面所示为具有额外中心体的一个人细胞中的一个过渡性多极纺锤体中间形态。图上所示微管为红色,中心体为绿色,染色体为白色。http://www.natureasia.com/zh-cn/nature/highlights/23560/
Nature 460, 278-282 (9 July 2009) | doi:10.1038/nature08136; Received 6 February 2009; Accepted 18 May 2009; Published online 7 June 2009
A mechanism linking extra centrosomes to chromosomal instability
Neil J. Ganem1, Susana A. Godinho1 & David Pellman1
Correspondence to: David Pellman1 Correspondence and requests for materials should be addressed to D.P. (Email: david_pellman@dfci.harvard.edu).
Chromosomal instability (CIN) is a hallmark of many tumours and correlates with the presence of extra centrosomes1, 2, 3, 4. However, a direct mechanistic link between extra centrosomes and CIN has not been established. It has been proposed that extra centrosomes generate CIN by promoting multipolar anaphase, a highly abnormal division that produces three or more aneuploid daughter cells. Here we use long-term live-cell imaging to demonstrate that cells with multiple centrosomes rarely undergo multipolar cell divisions, and the progeny of these divisions are typically inviable. Thus, multipolar divisions cannot explain observed rates of CIN. In contrast, we observe that CIN cells with extra centrosomes routinely undergo bipolar cell divisions, but display a significantly increased frequency of lagging chromosomes during anaphase. To define the mechanism underlying this mitotic defect, we generated cells that differ only in their centrosome number. We demonstrate that extra centrosomes alone are sufficient to promote chromosome missegregation during bipolar cell division. These segregation errors are a consequence of cells passing through a transient ‘multipolar spindle intermediate’ in which merotelic kinetochore–microtubule attachment errors accumulate before centrosome clustering and anaphase. These findings provide a direct mechanistic link between extra centrosomes and CIN, two common characteristics of solid tumours. We propose that this mechanism may be a common underlying cause of CIN in human cancer.
Controlling multipolar divisions in multi-centrosome cells
Micropatterns were used by Kwon et al. to demonstrate that interphase cell adhesion pattern and cell shape have a direct influence on multipolar divisions in MDA-231 cancer cells containing an excess number of centrosomes. Cells on Y and O shaped fibronectin patterns had a three-fold higher percentage of multipolar divisions versus the control (no patterns), whereas plating cells on a H shaped pattern suppressed 50% of multipolar divisions versus the control. Thus a bipolar pattern (the H) bias cells into bipolar mitoses and a multipolar pattern (the Y or the O) bias the cells into mutipolar mitoses.

(Figure from Kwon et al.)
|
|
Department of Pediatric Oncology, Dana-Farber Cancer Institute, Harvard Medical School, Boston, MA 02115, USA.
Multiple centrosomes in tumor cells create the potential for multipolar divisions that can lead to aneuploidy and cell death. Nevertheless, many cancer cells successfully divide because of mechanisms that suppress multipolar mitoses. A genome-wide RNAi screen in Drosophila S2 cells and a secondary analysis in cancer cells defined mechanisms that suppress multipolar mitoses. In addition to proteins that organize microtubules at the spindle poles, we identified novel roles for the spindle assembly checkpoint, cortical actin cytoskeleton, and cell adhesion. Using live cell imaging and fibronectin micropatterns, we found that interphase cell shape and adhesion pattern can determine the success of the subsequent mitosis in cells with extra centrosomes. These findings may identify cancer-selective therapeutic targets: HSET, a normally nonessential kinesin motor, was essential for the viability of certain extra centrosome-containing cancer cells. Thus, morphological features of cancer cells can be linked to unique genetic requirements for survival.
http://www.ncbi.nlm.nih.gov/pubmed/18662975
Multipolar spindles
Multipolar spindles are spindle formations characteristic of cancer cells. Spindle formation is mostly conducted by the aster of the centrosome which it forms around itself. In a mitotic cell wherever two asters convene the formation of a spindle occurs.[1]
Mitosis consists of two independent processes: the intra-chromosomal and the extra-chromosomal (formation of spindle) changes both of these being in total coordination of each other.

In cancer cells, it has been observed that the formation of the spindles comes before when com pared to the chromosomes. Due to this the prophase stage is briefer allowing metaphase to occur in advance. Chromosomes then are unable to reach the metaphase plate and are stranded behind. These chromosomes still have asters attached them and when met with other asters, multiple spindles are formed.[1]
CharactersticsCells with multipolar spindles are characterized by more than two centrosomes, usually four, and sometimes have a second metaphase plate. The multiple centrosomes segregate to opposite ends of the cell and the spindles attach to the chromosomes haphazardly. When anaphase occurs in these cells, the chromosomes are separated abnormally and results in aneuploidy of both daughter cells.[2] This can lead to loss of cell viability[3] and chromosomal instability.[4]
Presence in cancer cellsThe presence of multipolar spindles in cancer cells is one of many differences from normal cells which can be seen under a microscope. Cancer is defined by uncontrolled cell growth and malignant cells can undergo cell division with multipolar spindles because they can group multiple centrosomes into two spindles. These multipolar spindles are often assembled early in mitosis and rarely seen towards the later stages.

Research has shown possible causes of formation of multipolar spindles. A possible causes of multipolar spindle formation involve regulation of protein kinase family known as Aurora kinase.[5] Aurora kinase has two forms which are designated Aurora kinase A and Aurora kinase B.[6] These proteins play a key role in mitosis and are regulated by phosphorylation and degradation. Deregulation of these proteins can lead to multiple centrosome formation and aneuploidy.[5] In some human cancers, the expression and kinase activity of Aurora kinases have been up-regulated and has been looked into as a possible target for anti-cancer drugs.[5]
ReferencesArchiver|干细胞之家
( 吉ICP备2021004615号-3 )
GMT+8, 2024-5-7 00:20
Powered by Discuz! X1.5
© 2001-2010 Comsenz Inc.






